Research Article - Journal of Natural Product and Plant Resources ( 2018) Volume 8, Issue 3
Pearl millet [Pennisetum glaucum (L.) R. Br.] is a food cereal of limited importance in Benin. The conservation and exploitation of these resources are important for the food security of the populations involved in its production. Understanding the structure of its genetic diversity and identification on a molecular basis is an important goal for the support, preservation, and success of breeding programs. In this study, 42 varieties of millet collected in four agro-ecological zones of Benin already grouped into 3 morphological classes were evaluated using 12 microsatellite markers (SSRs). The analysis of the loci revealed a high level of polymorphism (100%), which detected a relatively large number of alleles (A=52) for an average of 4.3 per locus. Means of PIC, observed heterozygosity (Ho) and gene diversity (He) are respectively 0.63, 0.58 and 0.64. Typology of varieties by the UPGMA method using the SAHN procedure and the Principal Coordinate Analysis (PCoA) based on genetic distances showed the existence of three major genetic groups. The analysis of these results shows that farmers' practices have an impact on the genetic structuring of millet in Benin
Pearl millet [Pennisetum glaucum (L.) Br; Gramineae] is a highly cross-pollinated, diploid and annual cereal with seven chromosome pairs (2n=2x=14) [1,2]. Domesticated in West Africa [3-5], it has been broadcast around the world and has been the food of most countries in Africa and Asia. It is also a forage crop for some European countries. Millet is adapted to difficult environmental conditions, low capacity sterility where other cereals cannot withstand [4]. It is therefore ideal for the Sahel countries or countries of climatic conditions towards those of the Sahel.
In Benin, it is considered a secondary cereal with an annual production of 25182 tons [6]. It ranks 4th among the most important cereals after maize (Zea mays), sorghum (Sorghum bicolor) and rice (Oryza spp) with a yield of less than one ton of grain per hectare [6]. It is mainly used in human food and is very appreciated as a source of energy in certain ethnic groups in North-Benin [7]. Unlike maize and rice, millet does not have the opportunity for basic research that can significantly contribute to its improvement.
For any breeding program, the development of the best varieties must involve the value of seeds that are now considered sources of desirable genes or gene complexes. In Benin, prospecting and collecting forms of cultivated millet showed a great diversity of form in 1978 by Niandogo and Combes [8]. Despite this great diversity, no study has been established for its evaluation. Knowledge and evaluation of the genetic diversity of a crop are essential for the development of elite varieties adapted to the climatic conditions and local soil types of this crop. In most countries of the sub-region, to evaluate the genetic diversity of millet, many tests based on morphological and botanical traits have been conducted [9-15]. It has been shown that the evaluation of genetic diversity based on morphological characters conditions a limited number of genes [16]. It alone cannot ensure an accurate classification of the genetic divergence between accessions. However, molecular markers have been recognized as a powerful basis for assessing genetic variability and correlations among accessions of a given genetic resource [17].
Early on, millet diversity was assessed using iso-enzymatic loci [18,19], AFLP markers [20], RAPD markers [21] and RFLP markers [22,23] in several countries. Recently, new SSCP-SNP markers [24] and SSR [25,26] have been developed on millet varieties [5]. Among all these markers, SSRs have been declared the most reliable because of their multi-allelic property, their genomic specificity, their constant distribution, their high polymorphism, and their easy detection. In this study, their polymorphism was determined to evaluate the genetic diversity of local varieties of millet grown in Benin. It was specifically about (i) determining the genetic structuring of a collection of millet varieties using microsatellite markers; (ii) evaluate the impact of farming practices on this structuring.
Environment study and plant material
The study was carried out in thirty-two localities situated in four agro-ecological zones (Extreme North-Benin Zone (AEZ1), Northern Cotton Zone (AEZ2), Southern Borgou Food-producing Zone (AEZ3) and the Zone of West- Atacora (AEZ4)) of the North-Benin containing thirteen (13) different socio-cultural groups (Figure 1).
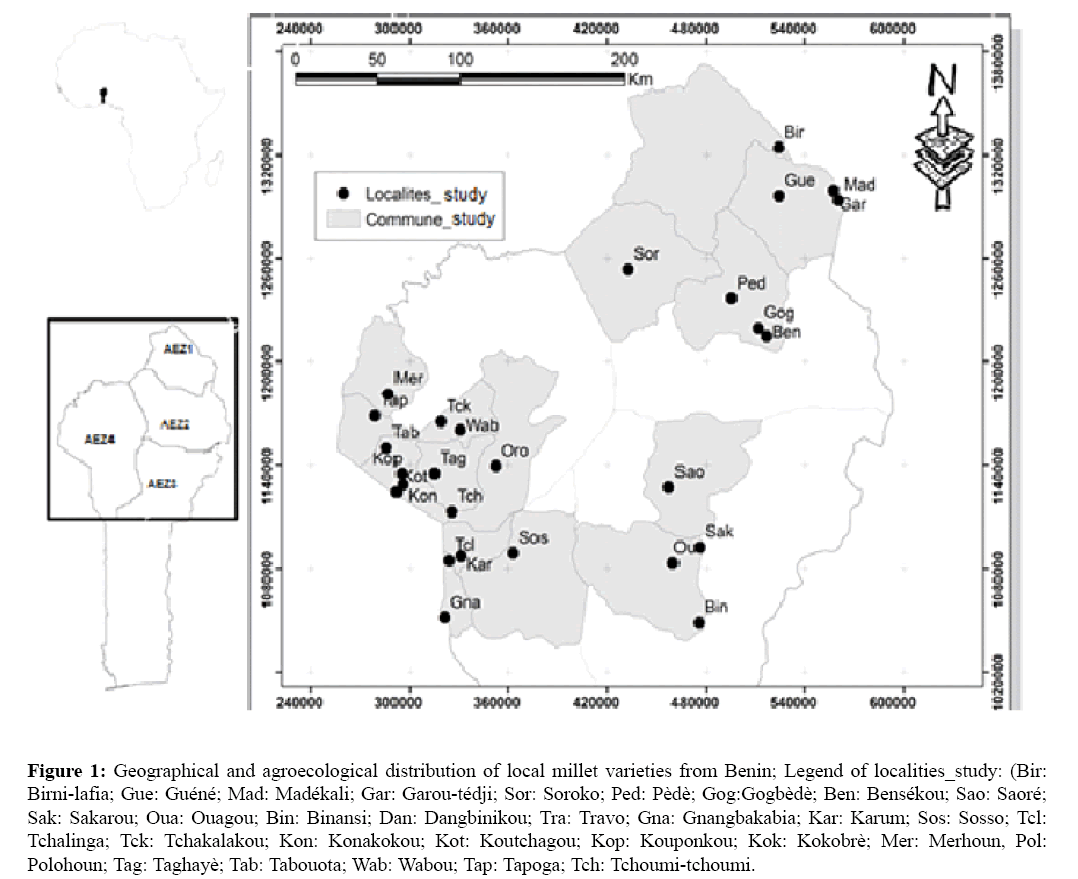
Figure 1: Geographical and agroecological distribution of local millet varieties from Benin; Legend of localities_study: (Bir: Birni-lafia; Gue: Guéné; Mad: Madékali; Gar: Garou-tédji; Sor: Soroko; Ped: Pèdè; Gog:Gogbèdè; Ben: Bensékou; Sao: Saoré; Sak: Sakarou; Oua: Ouagou; Bin: Binansi; Dan: Dangbinikou; Tra: Travo; Gna: Gnangbakabia; Kar: Karum; Sos: Sosso; Tcl: Tchalinga; Tck: Tchakalakou; Kon: Konakokou; Kot: Koutchagou; Kop: Kouponkou; Kok: Kokobrè; Mer: Merhoun, Pol: Polohoun; Tag: Taghayè; Tab: Tabouota; Wab: Wabou; Tap: Tapoga; Tch: Tchoumi-tchoumi.
The plant material studied includes 42 varieties of millet collected in December-January 2012-2013 and December- January 2013-2014 (Table 1). This collection was used for an agromorphological assessment of pearl millet varieties grown in Benin and allowed the determination of quantitative and qualitative traits, including the color of the grains used in our study [15].
| AEZs | Commune | Localities | Names of variety | Maturing cycle | Codes | Ethnic groups | Color of grains |
|---|---|---|---|---|---|---|---|
| AEZ4 | Natitingou | Taghayè | Yèyo mata | Late (4-6months) | Ytag | Otamari | Gray-brown |
| Tchoumi-Tchoumi | Ytt | Gray-dark | |||||
| Koutchagou | Ykt | Gray-dark | |||||
| (Koutagou) | |||||||
| Kounakokou | Ykn | Gray-brown | |||||
| Kouponkou | Ykp | Cream | |||||
| (Kouporgou) | |||||||
| Tabouota | Ytab | Ivory | |||||
| Toukountouna | Kokobrè | Yko | Gray-dark | ||||
| (Kokoko) | |||||||
| AEZ3 | Bembèrèkè | Dangbinikou | Yda | Mixture | |||
| (Dananzi) | |||||||
| Saoré | Ysa | Yellow | |||||
| Travo (Ouenou) | Ytr | Gray | |||||
| Sakarou | Ayo yantè | Late (5 months) | Asa | Niendé | Gray | ||
| Kouandé | Oroukayo | Yom mali | Late (4-5 months) | Yor | Natemba | Gray | |
| AEZ4 | Matéri | Polohoun | Itoura | Late (5 months) | Ipo | Berba | Gray |
| /Firihoun | |||||||
| Merhoun | Ime | Gray | |||||
| Toukountouna | Wabou | Sowawia | Late (4-5 months) | Swa | Wama | Gray | |
| Tchakalakou | Stc | Wama | Gray | ||||
| Ouaké | Tchalinga | Amala | Late (4 months) | Atc | Lokpa | Ivoire | |
| Djougou | Sosso | Amala Koupètè | Akps | Gray-dark | |||
| Sosso | Amala kouhloumè | Akhs | Gray | ||||
| Copargo | Karum | Na sowli | Early (3 months) | Nka | Tchila | Gray-dark | |
| Ouaké | Gnagbakabia | Maowairi ranéiri | Late (4 months) | Mgn | Peuhl | Ivory | |
| Cobly | Tapoga | Yotorika | Late (4-5 months) | Ytap | Kountèmba | Gray-Brown | |
| AEZ1 | Karimama | Birni-lafia | HainiKiré | Early (3 months) | Hbl | Dendi | Yellow |
| Malanville | Toumboutou | Hto | Yellow | ||||
| Madecali | Hma | Yellow | |||||
| Garou-Tédji | Hgt | Yellow | |||||
| Guéné | Hgu | Yellow | |||||
| Garou-Tédji | HainiKiré Dogo | Early (3 months) | Hdgt | Yellow | |||
| Karimama | Birni-lafia | Bandadabou | Early (2-3 months) | Ban | Gray | ||
| Malanville | Toumboutou | Somna | Late (4-5 months) | Sto | Yellow | ||
| Madecali | Sma | Yellow | |||||
| Garou-Tédji | Sgt | Yellow | |||||
| Guéné | Sgu | Yellow | |||||
| AEZ2 | Banikoara | Soroko | Gbè | Late (4-6months) | Gso* | Bariba | |
| Ouagou/Wagou | Gbè Kpika | Gou | Yellow | ||||
| AEZ3 | N'Dali | Binansi | Gbè Wonka | Gbp | Gray | ||
| Binansi | Gbè Wonka | Gbg | Gray | ||||
| AEZ2 | Kandi | Pèdè | Iyakou | Late (4 months) | Ipe* | Mokolé | Gray |
| Gogbèdè | Iyassè | Very early (2 months) | Igo | Gray | |||
| Bensékou | Wéagban | Late (4months) | Wbb | Bo | Gray-dark | ||
| Bensékou | Wéaguézénon | Wgb | Gray-dark | ||||
| Bensékou | Naaténon | Early (3months) | Nbe | Yellow |
Table 1: List of millet varieties collected by name and code, maturing cycle, ethnicity, locality, commune and AEZs.
The grains of each variety is planted in the greenhouse of the Laboratory of Genetics and Biotechnology of the University of Abomey-Calavi inside pots treated with carbofuran (insecticides) 72 Hours before sowing. One week later, the young leaves are taken for the extraction of the DNA.
DNA extraction
The extraction of DNA from the samples was carried out according to the protocol proposed by [27] using MATAB (Mixel Alkyl Trimethyl Ammonium Bromide), modified by [28] and adapted to various tropical crops. This protocol has been used successfully to several tropical plants such as Dioscorea spp. [28,29], Solenostemon rotundifolius [30], Sorghum bicolor [31]. About 0.2 g of young 8-day-old millet leaves are removed and then be crushed with 2 ml of Tris-EDTA sorbitol buffer (TES).
The crushed material is centrifuged at 10000 rpm during 10 min at 4°C. To the pellet obtained, 750 μl of 4% MATAB buffer preheated to 65°C are added. The whole is heated in a bain-marie at 65°C for 1 hour 30 min. Then, 750 μl of chloroform/isoamyl alcohol 24:1 is added to the mixture cooled in the ambient temperature and delicately agitated by turning for 2 min and then centrifuged at 10000 rpm during 15 min at 4°C. An equal volume of very cold isopropanol is added to the upper aqueous phase containing the DNA to precipitate the DNA. After centrifugation, the floating is carefully removed and the obtained ball is purified in 70° of ethanol, dried and then returned in sterile ultra-pure water. The DNA quality of the extracts has been evaluated by electrophoresis in a gel of agarose at 0,8%; Colored with bromide of ethidium (BET) and then visualized on a UV trans-illuminator. The extracted DNA was stored at -20°C. for further analysis.
Amplification by polymerase chain reaction (PCR)
Twelve (12) microsatellite markers revealed polymorphic in millet, identified by [32] and [5], tested by [33] have been used to characterize the genetic diversity of the varieties collected (Table 2). Dilutions of the DNA extracts have been carried out to obtain concentrations of the order of 10 ng/μl required for the amplification. The thermocycler used for amplification is Peltier-Effect Cycling (PTC-100TM) type.
| N° | Markers | Primer sequences 5’> 3’ F and R | Type of SSRs | PCR Products | Temperature of Hybridization |
|---|---|---|---|---|---|
| 1 | PSMP 2231 | TTGCCTGAAGACGTGCAATCGTCC | (TG)12GG(TA)4 | 229 | 52 |
| CTTAATGCGTCTAGAGAGTTAAGTTG | |||||
| 2 | PSMP 2232 | TGTTGTTGGGAGAGGGTATGAG | (TG)8 | 233 | 50 |
| CTCTCGCCATTCTTCAAGTTCA | |||||
| 3 | PSMP 2237 | TGGCCTTGGCCTTTCCACGCTT | (GT)8 | 233 | 56 |
| CAATCAGTCCGTAGTCCACACCCCA | |||||
| 4 | PSMP 2274 | CACCTAGACTCTACACAATGCAAC | (GA)13 | 265 | 50 |
| AATATCAAGTGATCCACCTCCCAA | |||||
| 5 | PSMP 2251 | TCAAACATAGATATGCCGTGCCTCC | (TG)6 | 162 | 54 |
| CAGCAAGTCGTGAGGTTCGGATA | |||||
| 6 | PSMP 2267 | GGAAGGCGTAGGGATCAATCTCAC | (GA)16 | 241 | 60 |
| ATCCACCCGACGAAGGAAACGA | |||||
| 7 | PGIRD 13 | CAGCAGCGAGAAGTTTAGCA | (AGC)8 | 250 | 60 |
| GCGTAGACGGCGTAGATGAT | |||||
| 8 | PGIRD 25 | CGGAGCTCCTATCATTCCAA | (GA)9 | 165 | 58 |
| GCAAGCCACAAGCCTATCTC | |||||
| 9 | PGIRD 43 | GTTCATGCAGCTTGGTTTCC | (GAT)6 | 100 | 54 |
| AGTGACCTGGGGTACAGACG | |||||
| 10 | PGIRD 50 | CTCTCGGTTTGACGGTTTGT | (TGT)6 | 120 | 50 |
| GGGGAAAACAAAGTTGCTCA | |||||
| 11 | PGIRD 57 | GGCCCCAAGTAACTTCCCTA | (AG)7 | 135 | 56 |
| TCAAGCTAGGGCCAATGTCT | |||||
| 12 | PGIRD 49 | AGCTCCTCGACGGAGAAAGT | (CGG)6 | 190 | 52 |
| GACGGTGTCGACGAAGATG |
Table 2: Characteristics of microsatellite millet markers used; Identified by [32] and [5] attested by [33].
The amplification cycle comprises pre-denaturation at 94°C for 5 min followed by 35 cycles, each consisting of denaturation at 94°C for 45 sec, of primer peer-specific temperature hybridization (50°C-60 °C) for 45 sec and elongation at 72°C for 1 min. A final incubation at 72°C for 30 min terminates the program. PCR products are checked on 2% agarose gel. A migration of the amplifiers by electrophoresis has been carried out on a 5% denaturing polyacrylamide gel. The migration time is from 1 h 30 min to 2 h. After migration, the revelation of the electrophoregrams is made with silver nitrate.
Data analysis
The genetic diversity parameters such as: number of alleles per locus (A), numbers of rare alleles (Ar) (<5% frequency), fairly frequent alleles (0,06<FA≤0,19) ; frequent alleles (0,20<FA≤0,50) and highly frequent alleles (0,51<FA≤0,98), allelic diversity, the number of observed heterozygosity (Ho) by [34] have been estimated. The Polymorphism Information Content (PIC) values have been calculated according to the algorithm of [35,36]: , with Fi and Fj less frequences of i and j alleles. The gene diversity or expected heterozygosity (He) has been calculated as , where n is the number of individuals and fi, the frequency of the i allele.
Based on the matrix (0,1), genetic distances of [34] are calculated between peers of local varieties. The distance matrix is then used to construct a dendrogram according to the UPGMA (Unweighted Pair-Method Method using the Arithmetic average) method and according to the SAHN (Sequential Agglomerative Hierarchical Nested method), software NTSYS version 2.21f [37,38]. In addition, to confirm the possible grouping of the analyzed varieties, the DCENTER and EIGEN procedures of this software are also used to perform a Principal Coordinate Analysis (PCoA) based on the same genetic distance matrix.
Genetic polymorphism revealed by microsatellite markers
The analysis of the SSR loci revealed a high rate of polymorphism (100%). The 12 loci used resulted in a total of 52 alleles (A=52). Of these 52 alleles detected, 5 alleles are rare (9.61%); 25 are moderately frequent (48.07%); 18 are frequent (34.62%) and 4 are highly frequent (7.69%). The number of rare alleles observed varies from 0 (PGIRD 43, PGIRD 50, PGIRD 49, PGIRD 25, PSMP 2232, PSMP 2267, PSMP 2231, PSMP 2251, PSMP 2237) to 2 (PGIRD 57). The number of alleles per locus varies from 2 (PGIRD 43, PGIRD 50) to 8 (PGIRD 57) with an average of 4.3 per locus (Table 3). The 12 SSR loci studied show a PIC ranging from 0.28 (PGIRD 50) to 0.85 (PGIRD 57) with an average of 0.63. A rate of heterozygosity (Ho) ranging from 0.07 (PSMP 2231) to 0.97 (PGIRD 49, PSMP 2267) with an average of 0.59 was observed. The gene diversity (He) ranges from 0.29 (PGIRD 50) to 0.86 (PGIRD 57) with an average of 0.64 (Table 3).
| Markers | A | 0,01< FA ≤ 0.05 | 0,06 < FA ≤ 0,19 | 0,20< FA ≤0,50 | 0,51< FA ≤ 0,98 | PIC | Ho | He |
|---|---|---|---|---|---|---|---|---|
| PSMP 2231 | 4 | 0 | 3 | 1 | 0 | 0.6 | 0.07 | 0.61 |
| PSMP 2232 | 7 | 1 | 5 | 1 | 0 | 0.8 | 0.92 | 0.81 |
| PSMP 2237 | 4 | 0 | 0 | 4 | 0 | 0.74 | 0.73 | 0.75 |
| PSMP 2274 | 4 | 1 | 2 | 0 | 1 | 0.46 | 0.29 | 0.47 |
| PSMP 2251 | 3 | 0 | 2 | 0 | 1 | 0.5 | 0.31 | 0.51 |
| PSMP 2267 | 5 | 0 | 2 | 3 | 0 | 0.77 | 0.97 | 0.78 |
| PGIRD 13 | 5 | 1 | 1 | 3 | 0 | 0.73 | 0.87 | 0.74 |
| PGIRD 25 | 4 | 0 | 2 | 2 | 0 | 0.7 | 0.75 | 0.71 |
| PGIRD 43 | 2 | 0 | 0 | 1 | 1 | 0.46 | 0.26 | 0.47 |
| PGIRD 50 | 2 | 0 | 1 | 0 | 1 | 0.28 | 0.24 | 0.29 |
| PGIRD 57 | 8 | 2 | 6 | 0 | 0 | 0.85 | 0.68 | 0.86 |
| PGIRD 49 | 4 | 0 | 1 | 3 | 0 | 0.71 | 0.97 | 0.71 |
| Means | 4.33 | 0.42 | 2.08 | 1.5 | 0.33 | 0.63 | 0.58 | 0.64 |
Table 3: Parameters of millet genetic diversity.
Structuring revealed by microsatellites
The estimated genetic distances between local millet varieties ranged from (D=0.05) between Gbp and Gbg at (D=0.99) between Hbl and Ykp, an average of 0.52. These genetic distances made it possible to construct a dendrogram based on the UPGMA method (Figure 2).
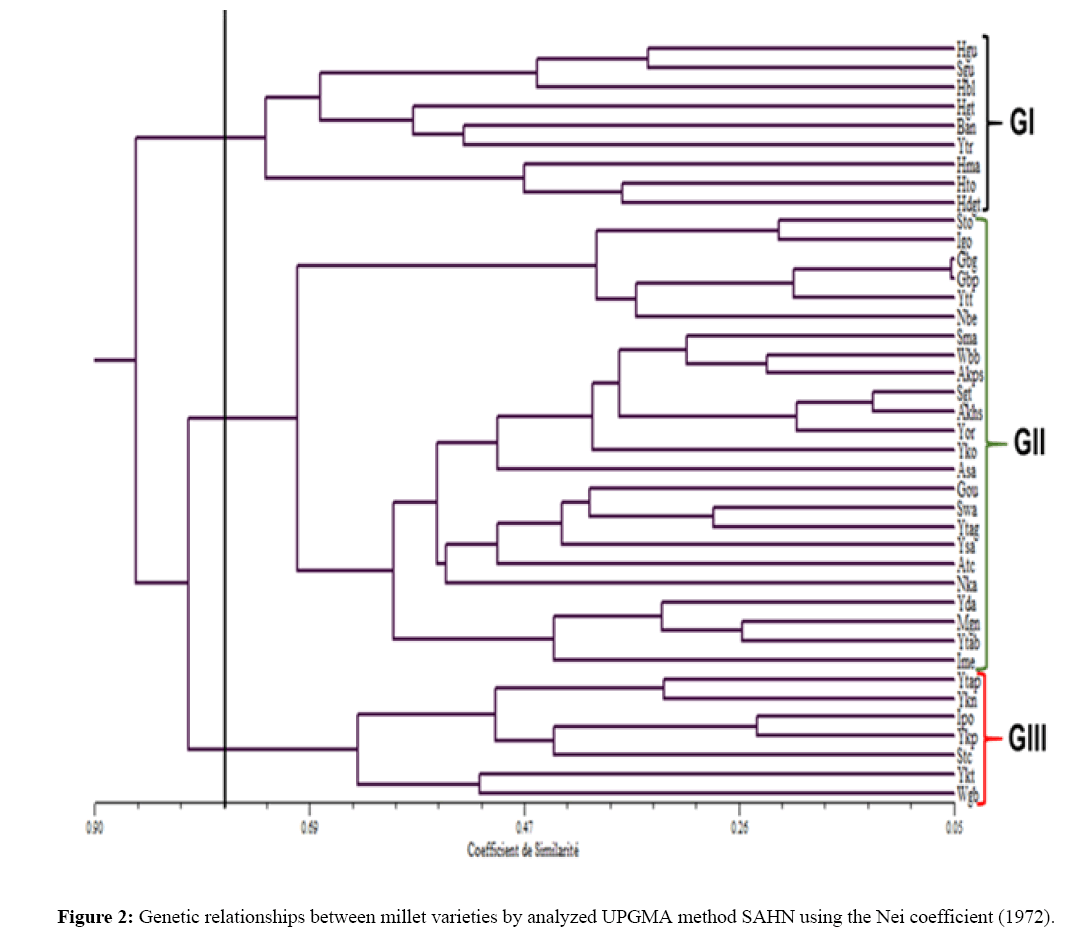
Figure 2: Genetic relationships between millet varieties by analyzed UPGMA method SAHN using the Nei coefficient (1972).
At the 77% similarity threshold, local varieties are organized into three major groups (GI GII GIII). The GI is composed of 9 varieties representing 22.50% of the local varieties analyzed, GII, 24 local varieties, ie 60%, and GIII, 7 local varieties, ie 17.50%.
The typological analysis of dendrogram shows that GI contains 77.78% of early variety (3 months) of the yellow color of hainikiré type (Hma, Hto, Hbl, Hgt, Hgu Hdgt), and Bandadabou (Ban) varieties from all of the AEZ1. This group includes two late varieties of the Somna (Sgu) type of AEZ1 and yeyo mata (Ytr) of AEZ3.
The GII consists of 87.50% of late varieties from AEZ1, AEZ2, AEZ3, and AEZ4 respectively 12.50%, 12.50%, 16.67% and 54.17%. These varieties vary in color of grain from Ivory (12.50%) to yellow (25%), gray 58.33% (gray, dark gray, brown gray) to the mixture of gray-white (4.16%). It includes three early varieties of AEZ2 (Igo, Nbe) and AEZ4 (Nka).
The GIII contains only late varieties (100%), mostly from AEZ4 (85.71%). They are gray color (85.71%) and cream (14.16%), Yeyo mata type (Ykn, Ykp, Ykt), Sowawia (Stc) and Yotorika (Ytap). It includes a local Wea type (Wgb) belonging to the dark-gray AEZ2.
From these three groups, we note that some varieties bearing the same local name, collected in two different localities and coming from the same AEZ belong to the same genetic group. This is the case of the local varieties Amala Koupètè (Akps) and Amala kouhloumè (Akhs) of the locality 'Sosso' and Amala kouhloumè (Atc) of the locality 'Tchalinga' collected in the AEZ4. Other varieties of the same AEZ, named in the same way, are found in different genetic groups. This is the case for the Itoura local varieties (Ime, Ipo) collected in two different localities of the AEZ4 are genetically different.
On the other hand, it has been observed that varieties bearing the same local names, coming from different agroecological zones and localities, have the same genetic base. These are the local varieties Yeyo mata of Kokobrè (Yko), Taghayè (Ytag), Tabouota (Ytab) and Tchoumi-tchoumi (Ytt) of the AEZ4 and Yeyo mata of Dangbinikou/ Dananzi (yda) and Saoré (Ysa) of AEZ3.
The distribution of genetic groups according to ethnic groups and AEZ shows that the local varieties of the GI are more specific to the Dendi ethnic groups in the AEZ1.
The local GII and GIII varieties did not show any specificity with regard to an ethnic group, but the local GIII varieties are mainly found in the ZAE4 whereas the GII varieties are found in all the AEZs (Table 4).
| AEZs | Ethnic groups | genetic groups | Total | ||
|---|---|---|---|---|---|
| GI | GII | GIII | |||
| 1 | Dendi* | 8* | 3 | 0 | 11 |
| 2 | Bariba, Bo*, Mokolé | 0 | 4 | 1 | 5 |
| 3 | Otamari, Bariba, Niendé, Natemba | 1 | 6 | 0 | 7 |
| 4 | Otamari*, Wama*, Berba*, Kountemba*, Lokpa, Peuhl, Tchila | 0 | 11 | 6* | 17 |
| Total | 9 | 24 | 7 | 40 | |
Table 4: Distribution of genetic molecular groups by ethnic group and AEZs ;* Majority in such ethnic group.
The Main Coordinate Analysis (PCoA) confirmed for a total variation of 51.13%, the classification of local varieties into three major groups with a slight approximation of certain local varieties of groups GI and GII. Axis 1 expressed 36.68% of the total variation and axis 2 the remaining 14.45% (Figure 3).
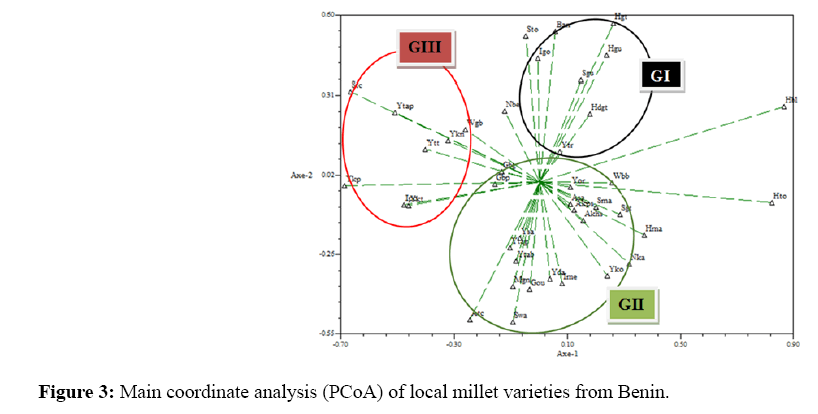
Figure 3: Main coordinate analysis (PCoA) of local millet varieties from Benin.
Genetic diversity and population structure
The effective use of locally adapted diversity is a major requirement for enhancing grain selection in breeding [39]. This study is the first to analyze the genetic diversity of local millet varieties in Benin using microsatellite markers. Of the 40 local varieties analyzed, the average gene diversity observed is (He=0.64). It is higher than that (He=0.62) reported by [40] on 115 R-line (Restorer) of millet and low (He=0.77) by [39] out of 214 millet accessions from Sudan. The difference may be due to the type of collection analysed. Thus, [40] used strains derived lines from the varieties whereas we used local varieties which according to [39] have a high level of heterozygosity and great diversity. Moreover, this observed difference compared to the results of [39] can be justified by the high number of markers having a large proportion of repeat dinucleotidic units which appears to be more variable than the long repeating units. The average heterozygosity observed (Ho=0.58) is relatively high given the size of the sample and the number of markers used in our study. This high level of observed heterozygosity can be attributed to the biological characteristic of millet, which is highly allogamic with a high rate of up to 85% [2,20].
Moreover, the average number of rare alleles (Ar=0.25) observed in this study is much lower than that (Ar=3.5) reported by [39] on millet. This can be explained by the fact that the local varieties used have a common ancestral origin with rare introduction during human migrations as the case of the local variety Bandadabou which despite its introduction, is neither morphologically nor genetically different from the Hainikiré varieties of the AEZ1 [7,14]. This may also be justified by the abandonment of certain local varieties by producers, such as the case of "Ignati" and "guéro" in the face of various constraints reported by producers [7,15].
The high level of gene diversity, the observed heterozygosity and the average number of alleles per locus observed in this study demonstrated the existence of a certain genetic difference between the local millet varieties analyzed. Similarly, the average genetic distance (0.52) obtained between the local varieties and the structured obtained from the dendrogram and PCoA confirmed this observation. The results showed that the local varieties of millet from Benin belong to 3 distinct genetic groups. This structuring in three groups confirmed the agromorphological study observed in the same collection, which also identifies three distinct classes, finally grouped into two morphological groups [14]. Some authors have identified 3 groups of millet varieties based on isozyme markers in Niger [4] and Burkina Faso [41]. Other genetic diversity studies based on SSR markers have identified a large number of genetic groups (5) but with collections with a large number of accessions or lines derived from millet varieties from several regions of the West and Central Africa [2,39].
Impact of farmer’s practices on genetic structuring
In our study, the main level of structuring of the genetic diversity identified seems to be based on the vegetative cycle which informs about the precocity of the local varieties G1 (Early) G2 (late with some early) and G3 (strict late). This level of structuring constituted partly the first criterion on which the local taxonomy of millet is based in all prospected areas [7] and then seems to agree with the conception of the producers by demonstrating clearly the influence of practices peasant management of the diversity on the millet genetic structuring. Nevertheless, the genetic structuring of local varieties on the molecular genetic basis testified of its importance by also showing the limits of the peasant classification. Some traditional names given by producers have not reflected the perfect identity of the genotype of individuals. Such observations have already been reported in the sub-region on the fonio [42] in Togo, rice [43] in Guinea, Sorghum [44] in Benin. In this study, it is the case of the presumed early and/or very early local varieties such as Naali (Nka) cultivated by Tchila (AEZ4), Naantenon (Nbe) by Bo (AEZ2), Iyassè by Mokolé (AEZ2) Which were found in the genotypic group GII. Let us remind here that the group GII is a group containing 87.5% of late varieties. It is very heterogeneous on the basis of the features putting out both the peasant identification and morphological classification (vegetative cycle, grain color, size of the candle) [7,14]. This group GII, together with the group GIII, seems to be closer to the big group of the late millet of West Africa [2,15,18] which according to [45] regroup both homogeneous and long short (homogeneous and heterogeneous) short-lived varieties. Also on the basis of breeds according to Brunken, GII and GIII seem very close? The great diversity of local names did not necessarily imply a great genetic diversity.
In addition, the presence of the late varieties Somna (Sgu) of AEZ1 and Yeyo mata (Ytr) varieties of AEZ3 in the genotypic group GI (early group) may be justified by the fact that in Benin, the low local diversity of millet within families could lead to, after several anthropogenic selections, a phenotypic homogeneity of local varieties. This homogeneity could create confusion at the level of the appellations (Somna case) or a false name due to the fact that the generic name of millet in the Otamari is Yeyo mata and there is no variant according to the producers (case of Yeyo). The presence of local varieties of all agroecological zones in the GII showed that there are grain exchanges between the agroecological zones studied.
Among the three genotypic groups obtained, the GI appears very homogeneous and more specific to the Dendi ethnic group (AEZ1). This result may be justified by the fact that AEZ1 is demonstrated to be an area where millet cultivation remains a very old practice [7]. Also its cohabitation with Niger and Nigeria (major producers of millet with known structuration) and its edapho-climatic characteristics (marked in part by the climate of the Sudano- Sahelian zone of West Africa conducive to culture would have enabled it to acquire this stability. The results obtained in this study of molecular genetic analyzes are very important for plant breeding programs of millet varieties adapted to the climatic conditions of the Agroecological Zones of Benin. They are also important for developing in situ conservation programs on local varieties of millet grown in Benin.
This study calls on to the microsatellite markers to clarify the diversity of the local varieties of the millet in Benin. It confirmed the efficacy and the potential of SSR markers in the evaluation of the genetic diversity of crops as it has been shown in the previous studies. It explored at the molecular genetic level the variability within the accessions of cultivated millet and showed the existence of a great genetic difference between the local varieties of millet analyzed. Besides, the study allowed to obtain three distinct genotypic groups, two of which seem to be very homogeneous (GI and GIII) and one heterogeneous (GII). These results have highlighted the impact of certain peasant practices on the genetic structure of millet. They are globally significant for any program of genetic improvement of this neglected speculation and can enable to define effective strategies of conservation in-situ of the genetic resources of the millet cultivated in Benin.
At the end of this study, we would like to sincerely thank those who contributed directly or indirectly to the realization of this work. We thank Dr. Valère SALAKO and the staff of the Laboratory of Molecular Genetics and Genome Analysis, especially the technicians Germaine AVOHOU and Jonas ALLOWANOU.